
Testicule
Cele testiculele [ Hodn ] sau (mai rar) sau HODE [ hoːdə ] (prin Orientul ridicat german HODE și veche germană de Hodo a indo - * skeu (t) sau "capac, capac") , testicul (din latină testiculus , diminutivul testicul "martor [al virilității ], testicul", plural: testicule ; greaca veche ὄρχις orchis ), din punct de vedere tehnic, testicul și testicul , este un organ sexual masculin pereche, intern, al multor animale de țesut care se reproduc sexual . La fel ca ovarele indivizilor feminini, aparține gonadelor ( gonadelor ) și produce filamentele seminale ( spermatozoizii ). În plus, în testicule se formează hormoni sexuali masculini ( androgeni ), în special testosteronul . La vertebrate, testiculele se formează embrionar în cavitatea abdominală, dar la majoritatea mamiferelor migrează în scrot ( scrot ).

anatomie
Mamifere
Dimensiune și locație

Testiculul uman are aproximativ o formă de prună , cântărește aproximativ 20 de grame și are un volum mediu de 20-25 ml. Lungimea medie este de 5 cm, grosimea de aproximativ 3 cm. Testiculele nu se dezvoltă până la dimensiunea maximă până la pubertate și ating dimensiunea maximă în deceniul 4 de viață. Odată cu vârsta, dimensiunea testiculului scade din nou. Volumul testiculului oferă informații despre starea funcțională a testiculului, printre altele. Dacă volumul testiculului este sub 8 ml, se poate presupune că producția de spermă funcționează doar într-o măsură limitată sau deloc. Testosteronul, pe de altă parte, este uneori încă produs până la un volum de 1,5 ml; sub aceasta, testiculul este de obicei disfuncțional.
La mamifere , forma testiculului variază de la rotunjită la ovoidă. Există diferențe clare de mărime, dar nu există o relație strânsă cu greutatea corporală. Cele mai mari testicule din lumea animalelor au balene drepte sudice , fiecare cu 500 kg reprezintă 2% din greutatea corporală. Rozătoarele , oile (până la 300 g fiecare) și porcii domestici (până la 750 g fiecare) au testicule relativ mari, în timp ce prădătorii sunt relativ mici . La animalele cu o perioadă sezonieră în reproducere, mărimea testiculului este, de asemenea, supusă fluctuațiilor sezoniere, testiculele sunt semnificativ mai mari în sezonul de împerechere decât în perioada de repaus.
La majoritatea mamiferelor, ambele testicule sunt la indivizi maturi sexual din scrot ( scrot ) sau saci de piele asemănători cu scrotul. Testiculele apar în cavitatea abdominală , dar migrează prin canalul inghinal în scrot în timpul nașterii, la rozătoare până la pubertate . Acest proces se numește descendență testiculară ( Descensus testis ). La unele mamifere (de exemplu , hamsteri , lilieci ) există o coborâre testiculară sezonieră, iar testiculele sunt doar în afara cavității abdominale în timpul sezonului de împerechere. Cu toate acestea, în cadrul mamiferelor există câteva grupuri de animale în care testiculele rămân în general în cavitatea abdominală, așa-numita testiconda . Testiculele pot rămâne la locul sistemului (ca la elefanți ) sau pot coborî, dar rămân în cavitatea abdominală (de exemplu la balene , vezi și tabelul).
| Testiconda | ||
| Fără coborâre testiculară | Coborâre incompletă a testiculelor | Coborâre testiculară sezonieră |
| Monotreme , crizocloride , scorpii , Igeltenreks , hidrele , elefanți , vaci de mare , lenea cu trei degete , anteaters | Balene , armadillo | Alunițe , gărgărițe , varci , șopârle , lilieci , niște rozătoare |
Structura anatomică
Structura anatomică externă a testiculului se bazează pe epididimul care se învecinează și se fuzionează cu acesta . Secțiunea testiculului care indică capul epididimului se numește capătul capului ( Extremitas capitata ), secțiunea care indică coada epididimului se numește capătul cozii ( Extremitas caudata ). La capătul cozii există adesea un rudiment nefuncțional, în formă de neg, al așa-numitului canal Müller , cunoscut sub numele de apendicele testicular ( apendicele testicul , o hidatidă Morgagni ). Marginea orientată către epididim este marginea epididimului ( Margo epididymalis ), opusă acestuia este marginea liberă ( Margo liber ). În plus, se pot distinge o suprafață orientată spre centru ( Facies medialis ) și o suprafață orientată spre exterior ( Facies lateralis ).
Cei testiculele coboară într - o umflătură în peritoneu și interior trunchiul fascia ( se face referire aici ca spermatica INTERNA fascine ), a vaginalis apofiza . Procesul vaginului este una dintre tecile testiculare din interiorul scrotului. Partea peritoneului din această protuberanță se numește pielea vaginală ( tunica vaginalis testicul ). Căptușește interiorul scrotului (așa-numita foaie de perete, lamina parietalis sau periorchium ), apoi se rotește în interior ca o lamelă dublă și acoperă testiculele ca o foaie intestinală ( lamina visceralis sau epiorchium ). Există un spațiu foarte îngust între cele două frunze, cavum vaginal , care asigură că testiculul se poate mișca în scrot. Punctul de legătură dintre cele două frunze este testicul cros ( mesorchium ), care este utilizat pentru a atașa testiculul la scrot. Testiculul este, de asemenea, la capătul cozii cu o bandă scurtă conectată la epididim (banda testiculului propriu, ligamentul testicul propriu ). Acest lucru continuă de la coada epididimală ca ligament de coadă epididimală ( ligamentum caudae epididymidis ) și atașează suplimentar testiculele indirect la podeaua scrotului. Mușchiul de ridicare a scrotului ( Musculus cremaster ) se atașează, de asemenea, la procesul vaginal, deoarece un dispozitiv de protecție trage testiculele mai aproape de peretele abdominal atunci când este atins sau rece. La rozătoare și mamifere cu descendență testiculară sezonieră, rareori și la indivizi individuali ai altor mamifere, mușchiul poate trage testiculul complet înapoi în cavitatea abdominală (" testiculul pendulului ").
O capsulă groasă de țesut conjunctiv albicios , tunica albuginea, se află direct sub acoperirea peritoneului testiculului . Asigură rezistența mecanică a organului și menține o anumită presiune internă. Din această capsulă septuri trageți în interior și subdivideți testiculele în lobuli testiculari ( lobuli testicul ). Testiculul bărbatului are aproximativ 350 de lobuli testiculari. Septa formează, de asemenea, un corp de țesut conjunctiv, mediastin testicul , care în anatomia umană este numit și corpus highmori .
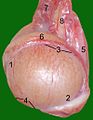
Testicule, epididim și cordonul spermatic al unei pisici de sex masculin :
1 capăt
2 capăt 2 coadă
3
marginea epididimului 4 margine liberă
5 mezenter testicular
6 epididim
7 plexul arterei și venei testiculare
8 canal spermatic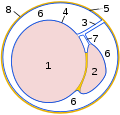
Secțiune transversală prin procesul vaginal:
1 testicul
2 epididim
3
mezenter testicular 4 foaie de organ a membranei vaginale
( epiorhiu )
5 foaie de perete a pielii vaginale
( periorhiu )
6 cavum vaginal
7 mezenter epididim
8 fascia spermatică internă
Secțiunea prin testiculul unui taur:
(vasele de sânge injectate cu gelatină roșie )
1 parenchim testicular
2 mediastin testicul
3 tunica albuginea
4 coada
epididimului 5 capul epididimului
6 cordonul spermatic cu
tendril convolut al arterei testiculare



Vasele și nervii
.jpg)
Testiculul este alimentat cu sânge prin artera testiculară ( artera testiculară ). Apare, corespunzător localizării atașamentului embrionar al testiculului (vezi mai jos), direct în spatele arterei renale, direct din aorta abdominală din regiunea lombară. La animalele cu descendență testiculară, artera testiculară trebuie să se lungească în mod corespunzător și rulează de-a lungul spatelui peretelui abdominal într-un mezenter scurt ( mesorchium proximal ) până la canalul inghinal . În afara cavității abdominale, acesta intră în cordonul spermatic . Aici se află în înfășurări spiralate strânse, așa-numita tendrilă convolută . La tauri , de exemplu, o secțiune arterială lungă de doi metri este găzduită pe o lungime a cordonului spermatic de 13 cm. Pachetul de tendril este înfășurat în plexul tendrilului ( Plexus pampiniformis ) al venei testiculare ( Vena testicularis ). Acest lucru creează o zonă mare de contact între sângele de intrare și de ieșire, care acționează ca un schimbător de căldură . Temperatura din scrot este cu câteva grade sub temperatura internă a corpului, ceea ce este esențial pentru formarea spermei fertile la mamiferele cu descendență testiculară. Sângele care intră în artă este răcit prin acest aranjament prin sângele care iese din venă.
Artera testiculară rulează de-a lungul marginii epididimului până la capătul cozii testiculului. De acolo se deplasează în interiorul capsulei testiculare la marginea liberă la majoritatea mamiferelor ( rumegătoarele , de exemplu, fac o excepție ) înapoi la capătul capului. Ramurile sale șerpuiesc în tunica albuginea peste suprafețele laterale și trec peste septurile testiculare din interior către testicul mediastin și de acolo centrifugal înapoi la tubulii seminiferi, în jurul cărora formează o rețea capilară .
Inervarea testiculului este mediată de simpatic , parte a sistemului nervos autonom . În fibrele nervoase provin din trunchiul regiunii lombare și, ambalaj în jurul arterei testiculare ca o împletitură ( plexul testicular , sinonim: nervul spermatic superioare ), se extind la testiculele. Un al doilea grup de fibre nervoase trece de la ganglionii încrucișați ai trunchiului cu ductul spermatic ( ductus deferens ) până la testicule ( plexus deferențial , sin. Nervus spermaticus inferior ). Cele eferente fibrele nervoase ajung in principal vasele de sânge și astfel reglează fluxul sanguin și temperatura testiculului. Participarea la controlul fin al formării spermei , transportului de spermă și producției de hormoni la testiculele unor mamifere este în prezent discutată, dar acest control se efectuează în principal prin intermediul hormonilor . La corpurile celulare ale acestor fibre nervoase care transporta informații către sistemul nervos central ( visceroafferences ) sunt situate în rădăcină dorsală ganglionilor din regiunea lombară. Acestea transmit senzații de durere ( dureri intestinale), dar o mare parte a sensibilității ridicate a testiculului la durere este transmisă prin fibrele nervoase sensibile ale tecilor testiculare (ramuri ale nervului genitofemoral ). Sensibilitatea ridicată la stimuli la atingere face din testicule și scrot o zonă erogenă . Pe de altă parte, marea sensibilitate la durere este exploatată și în BDSM ( tortură cu cocoș și mingi ) și tortură ; Strângerea testiculelor poate duce la șoc . O nevralgie a nervului genitofemoral, care poate apărea, de exemplu, după corecția chirurgicală a unei hernii inghinale , se manifestă prin durerea testiculară.
La vasele limfatice ale testiculului rula împreună cu venele testiculare la lombare ganglioni limfatici ( lumbales Lymphonodi ) înapoi a aortei, la mamifere domestice , de asemenea , la iliace ganglionii limfatici ( Lymphonodi iliaci mediales ) la ramura aortic. În cancerul testicular, metastazele pot apărea în acești ganglioni limfatici , care sunt localizați în spațiul retroperitoneal al abdomenului .
Acorduri rămase
În toate celelalte cordate , testiculele se află în cavitatea corpului și sub rinichi.
La scullless nu există testicul compact, gonadele sunt încă segmentate, în Asymmetron este prezent doar testiculul potrivit. La hagfish , testiculul alungit se formează, de asemenea, doar pe o parte, sub forma unei benzi lobate. Cele Gurile rotunde au împerecheat testicule care se extind pe întreaga lungime a cavității corpului.
În cadrul peștilor cartilaginoși , numai în rechinul guler , rechinul cel mai primit din punct de vedere anatomic și morfologic, întregul sistem testicular se dezvoltă într-o gonadă alungită, la ceilalți reprezentanți, ca și la celelalte vertebrate , doar la secțiunea mijlocie (vezi mai jos). Testiculele la rechini reale sunt , de asemenea , alungite, iar cele ale razelor sunt scurte și plate. La câinele și razele electrice , testiculele sunt departe în față, în apropierea pericardului , altfel în secțiunea centrală sau posterioară a trunchiului. La majoritatea peștilor osoși , testiculele sunt alungite. La peștii osoși reali , aceștia sunt situați sub rinichi și vezica înotătoare și sunt atașați printr-un mezorchiu. La unele specii de biban , ambele testicule sunt fuzionate împreună la capătul posterior. Tunica albuginea subțire este ocazional pigmentată în pești osoși . În același timp, țesutul ovarian poate apărea în testiculele peștilor (hermafrodit, ovotestis ). Saw bass și dorad sunt hermafrodite , deci au atât testicule, cât și ovare și pot schimba sexul pe parcursul vieții lor.
Amfibienii au testicule alungite ( amfibieni de coadă , amfibieni care se târăsc ) sau rotunjite ( amfibieni de broască ). Acestea sunt atașate de peretele trunchiului sau de rinichiul urnal printr-un mezorhiu . Un corp de grăsime distinct este format medial (în direcția planului median ) al testiculului. În salamandre , mai multe departamente testiculare sunt fuzionate într-un singur lob, numărul departamentelor crește odată cu vârsta. La amfibieni, tractul urinar și cel seminal încep să se separe. Canalele epididimale ( ductuli epididymidis ) se deschid în canalul Wolff doar cu puțin înainte de cloacă . Broaștele masculine sunt hermafrodite. În fața testiculelor se află organul licitator , un ovar primitiv, care se dezvoltă din anlajul gonadal anterior .
La reptile , testiculele sunt în fața și sub rinichi în imediata apropiere a glandelor suprarenale . Testiculele sunt ovale, mai rotunjite la broaște țestoase și alungite la șerpi . Epididimul este medial (spre centrul corpului) testiculului respectiv. Cadrul țesutului conjunctiv al testiculului este slab dezvoltat.
Și la păsări , testiculele se află în fața rinichilor pe glandele suprarenale. Spre deosebire de ovar, care este doar unilateral la păsări, testiculele sunt întotdeauna împerecheate. Mărimea testiculului prezintă cele mai mari fluctuații sezoniere în cadrul vertebratelor; crește, de exemplu, la păsările passeriforme de până la 800 de ori în timpul sezonului de împerechere și scade din nou în timpul somnului de împerechere. Într-o casă drakes testiculele sunt în sezonul de împerechere de aproximativ 8 cm lungime și 4,5 cm lățime. Structura țesutului conjunctiv al testiculului este ușor dezvoltată la păsări, tunica albuginea este subțire și nu există testicul mediastin . Endoscopica examinarea testiculului utilizat pentru a juca un rol major în determinarea sexului speciilor care prezintă nici un dimorfism sexual , dar astăzi a fost înlocuită în mare măsură cu biologice moleculare metode.
Nevertebrate
Reproducerea asexuală și sexuală are loc în cadrul cnidarienilor . În timpul reproducerii sexuale, celulele germinale sunt eliberate în apa înconjurătoare sau în spațiul gastric prin spargerea epiteliului . Meduzele coastei sunt în general hermafrodite și au testicule și ovare încorporate în mezogloea sub trombocite asemănătoare pieptenilor („coaste”) .
Guri primare (protostomie)
Majoritatea viermilor plati (trombocite) sunt hermafroditi. Nu au o cavitate corporală ( celom ), testiculele se află adiacente ovarelor într-un spațiu de țesut conjunctiv ( interstitiu ) din interiorul animalului, în cazul viermilor din fiecare membru al teniei ( proglotid ). Unele platelminții ( Catenulida și Acoelomorpha ) nu au gonade. De asemenea , părul de burtă (Gastrotricha) nu are celom, speciile de apă dulce se reproduc asexual prin partenogeneză . Reprezentanții apei sărate sunt hermafrodite, dar numai gonadele unui sex sunt active funcțional. Celulele germinale sunt eliberate prin grămezi de spermă . Rotiferele ( Rotatoria ) au un pseudocoelom cu testicule împerecheate sau nepereche, la unele specii apar așa-numiții masculi pitici , la alții nu există bărbați, reproducerea are loc apoi prin generarea virgină. Cycliophora se reproduc asexual în așa-numita etapă de hrănire. În reproducerea sexuală apar bărbați pitici. Au două „testicule” externe și un organ copulator lângă discul adeziv. În cazul viermilor zgârieturi (Acanthocephala), un așa-numit ligament genital străbate corpul, pe treimea inferioară a căreia sunt localizate cele două testicule. Un canal de spermă trece de la fiecare testicul la penis.
Priapulida și animale corset (Loricifera) au asociat testiculelor , care sunt unite cu protonephridia pentru a forma un aparat urinar și sexual . La viermi rotunzi (Nematoda) există atât specii de sex separat, cât și hermafrodite. Testiculul alungit se află sub intestin și, de asemenea, se deschide în el (vezi fig.). Viermii cu coarde (Nematomorpha) au testicule pereche în pseudocoelom. Tardigradele (Tardigrada) sunt sexe separate. În timp ce cavitatea corpului reprezintă un pseudocoelom, există un adevărat celom în jurul testiculelor nepereche. Colibus (onychophora) au asociat testiculele care se deschid într - un vas comun deferent prin conducte. La artropode (Arthropoda: milipede , insecte , crustacee și chelicerate ) masculii au testicule împerecheate în pseudocoelul plin de sânge ( hemocoel ) al abdomenului sau al segmentelor posterioare ale trunchiului. Există și hermafrodite în unele artropode.
Cei anelide au asociat testiculele. Viermii de centură sunt în majoritate hermafroditi, dar se fertilizează reciproc. Testiculele sunt localizate în celom , la râme în segmentele 10 și 11 ale corpului și canalele deferente se deschid spre exterior în segmentul 15. Multi-peri , pe de altă parte, sunt în mare parte unisexuali, iar reprezentanții bărbaților au testicule în fiecare segment al corpului. La moluște (Mollusca) există hermafrodite, gonade hermafrodite ( Ovotestis ) și forme sexuale separate. Celomul este redus la două cavități în jurul gonadelor ( gonadocoel ) și în jurul inimii ( pericard ) din spatele acestuia . La armpoduri (Brachiopoda), testiculele se află în celom (mai exact în metacoel ), gametii sunt derivați prin metanefridie .
Neumünder (Deuterostomia)
În echinoderme (Echinodermata), balenele de mare au un singur testicul, în arici și balene de mare testiculele umple aproape întregul metacoel, în crinii de mare (trei până la cinci gonade) și stelele de mare (două pe braț) întindeți-vă în brațe și deschideți-le între ele Brațele cu câte o deschidere genitală.

Organele interne ale unui pește osos:
7 testicule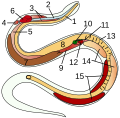
Organele interne ale unui șarpe:
14 testicule
Locul cocoșului domestic:
(cu excepția organelor digestive)
3 testicule
Viermi rotunzi:
5 testicule
Planul unei păstăi cu gonade în metacoel




Structura și funcția fină la vertebrate
Lobulii testiculari conțin fiecare câte doi până la patru tubuli seminiferi înfășurați ( tubuli seminiferi contorti s. Convoluti ), care reprezintă parenchimul testicular . Acestea au aproximativ 50 până la 80 cm lungime și 150 până la 300 um în diametru. Peretele său este format dintr-o coajă de țesut conjunctiv cu miofibroblaste contractile , o membrană bazală și epiteliul germinal ( epitelium spermatogenicum ).
Acest epiteliu este format din spermă sau celule germinale ( Cellulae spermatogenicae ) și celule Sertoli . Celulele spermatozoizilor sunt formate din celulele germinale ( spermatogeneza ). Deoarece spermatogeneza este cea mai importantă sarcină a testiculului, celulele germinale sunt de asemenea prezente cel mai frecvent în testicule în ceea ce privește cantitatea. În timpul formării spermei, etapele succesive de dezvoltare ale celulelor germinale ( spermatogonia → spermatocite → spermatide → spermatozoizi) sunt transportate treptat către lumen . Formarea spermei durează între 35 (șoarece, porc) și 64 de zile (uman), dar este necesară o maturare suplimentară în epididim, astfel încât sperma să poată fi fertilizată. Aceasta durează o săptămână la majoritatea mamiferelor și de la 8 la 17 zile la oameni. La bărbați, aproximativ 200 până la 300 de milioane de spermatozoizi sunt eliberați din epididim per ejaculare . Cu o ejaculare mai frecventă, cantitatea de spermă scade, deoarece capacitatea zilnică de producție a spermei este limitată. Depinde de masa testiculului și de numărul de celule Sertoli și este între 45 și 200 de milioane de spermatozoizi pe zi la bărbați.
A doua componentă importantă a tubulilor seminiferi sunt celulele Sertoli ( Epitheliocyti sustentantes ). Acestea au aproximativ 70 până la 80 µm lungime și rulează radial prin întregul epiteliu germinal până la lumen. Celulele Sertoli au o funcție de susținere și îngrijire pentru celulele spermatozoizilor, ele hrănesc celulele spermatozoizilor, asigură mediul lor hormonal corect și folosesc mișcările plasmatice pentru a le transporta la lumen. În plus, celulele Sertoli fagocitează celulele de spermă degenerate și resturile celulare care apar în timpul dezvoltării spermei. Celulele Sertoli sunt activate de hormonul foliculostimulant al hipofizei (FSH) controlat, distribuția acestuia asupra formării influenței hormonului inhibină B. În plus, ele secretă proteina de legare a androgenilor , hormonul anti- Müllerian și un lichid seminal bogat în potasiu .
Celulele Sertoli au numeroase procese care înconjoară celulele germinale. Aceste procese celulare se conectează la baza tubulilor seminiferi prin joncțiuni strânse cu cele ale celulelor Sertoli învecinate și formează astfel așa-numita barieră hemato-testiculară . Acest termen este de fapt înșelător, deoarece această barieră nu se află între sânge și țesutul testicular, ci se desfășoară între spermatogonia și spermatocitele, împărțind astfel tubulii testiculari într-o manieră circulară într-un compartiment bazal și într-un compartiment orientat spre lumen (adluminal) . Bariera sângelui-testicul este impermeabilă pentru majoritatea proteinelor și protejează sperma de mutageni și de propriile mijloace de apărare ale corpului . Aceasta din urmă este necesară deoarece primele celule de spermă apar doar după imprimarea limfocitelor (vezi autotoleranță ), astfel încât sistemul imunitar le-ar considera ca fiind celule străine. Cu toate acestea, citokinele antiinflamatorii , a căror eliberare este probabil dependentă de androgen și celulele sistemului imunitar din testicule ( celule dendritice , macrofage ) joacă, de asemenea, un rol în protejarea împotriva reacțiilor autoimune .
La multe vertebrate, tubulii seminiferi complicati se contopesc într-un tubul seminifer scurt drept ( tubulus seminifer rectus ) la ambele capete . Tubii drepți sunt căptușiți de un epiteliu cu un singur strat și se deschid într-un sistem de canal în mediastin, rețeaua testiculară ( rete testis ). Sistemul canalic al rețelei testiculare este, de asemenea, în mare parte căptușit de un epiteliu cu un singur strat (cu două straturi la bovine). Cu toate acestea , la bărbați, armăsari și rozătoare, rețeaua testiculară este predominant la periferia testiculelor („rete extratesticulară”). Câteva ductuli efemerante testiculare șerpuitoare rulează din rețeaua testiculară în capul epididimal și se unesc acolo pentru a forma canalul epididimal. La mamifere există aproximativ 15 ductule eferente, numărul variază în rândul vertebratelor între una (de exemplu, raze ) și 32 (de exemplu, axolotl ).
Țesutul dintre tubii seminiferi complicati este numit interstitiu . La majoritatea vertebratelor, acesta reprezintă aproximativ 10 până la 20% din țesutul testicular, în cazuri extreme, cum ar fi lemnul, aproape 70%. Pe lângă țesutul conjunctiv, vasele de sânge și fibrele nervoase , interstițiul conține și celule Leydig ( Endocrinocyti interstitiales ). Prin contacte speciale de celule ( joncțiuni gap ), ele formează asociații de celule interconectate, așa-numitele sincitii funcționale . În funcție de hormonul luteinizant (LH), celulele Leydig produc hormoni sexuali masculini (androgeni precum testosteronul și androstanolona ), precum și oxitocina , care promovează motilitatea tubulilor seminiferi. Testiculul este astfel și un organ endocrin . Testosteronul face ca spermatidele să se maturizeze în tubulii seminiferi. Pentru a trece prin bariera hemato-testiculară până la locul său de acțiune, are nevoie de proteina de legare a androgenilor a celulelor Sertoli. Androgenii au, de asemenea, o varietate de efecte în organism, inclusiv promovarea dezvoltării caracteristicilor sexuale secundare , având un efect anabolic și controlul comportamentului sexual . În plus, celulele Leydig produc numeroase alte peptide active hormonal care acționează asupra celulelor vecine ( paracrine ) sau asupra celulei Leydig în sine ( autocrine ) care le compune .

Imagine histologică a parenchimului testicular al unui mistreț:
1 lumen al unui tubul seminifer complicat
2 spermatide
3 spermatocite
4 spermatogonia
5 celule Sertoli
6 miofibroblaste
7 celule Leydig
8 capilare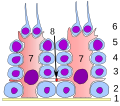
Epiteliul germinativ al tubului seminifer :
1 membrană bazală
2 spermatogonie
3 spermatocite ordinul 1
4 spermatocite ordinul 2
5 spermatide
6 spermatide mature
7 celulă Sertoli
8 joncțiune strânsă
(barieră sânge-testicul)
Schema structurii interne a testiculului și a epididimului :
1 tunica albuginea
2 septuri
3 lobi testiculari
4 mediastin testicul
5 tubuli seminiferi convoluți
6 tubuli seminiferi drepți
7 rete testicul
8 ductuli eferenți testicul
9 epididim
10 începutul canalului deferent



Control hormonal

Controlul hormonal al testiculului se efectuează prin atacuri și atacuri, de către celulele nervoase din eminența mediană din hipotalamusul format din gonadoliberină (GnRH). GnRH nu acționează direct asupra testiculelor, ci stimulează formarea hormonilor LH și FSH în hipofiza anterioară . Eliberarea acestor hormoni este, de asemenea, controlată de testiculele înseși printr-un mecanism de feedback negativ: secreția FSH este inhibată de inhibina B produsă de celulele Sertoli, în timp ce secreția GnRH este inhibată de testosteronul produs de celulele Leydig . Fluctuațiile sezoniere ale mărimii și activității testiculelor la multe animale sunt mediate de suprimarea secreției de GnRH în timpul odihnei sub influența lungimii zilei. Mecanismul exact nu este încă cunoscut în detaliu: la mamifere, opioizii , neuronii dopaminergici și melatonina sunt probabil implicați în această buclă de control, la păsări și hormoni tiroidieni .
LH se leagă de un receptor de membrană al celulelor Leydig și astfel induce sinteza androgenilor . În acest caz, colesterolul este treptat, printre altele, pregnenolon și progesteron , transformat în testosteron, utilizând două căi sintetice diferite (Δ4 și Δ5). Efectul LH asupra celulelor Leydig este potențat de prolactină , dar cu o supraproducție de prolactină apare prin reglarea descendentă a receptorilor LH la inhibarea sintezei testosteronului. LH poate induce, de asemenea, formarea de androgeni în cortexul suprarenal ; dehidroepiandrosteronul format acolo ajunge la testicule prin sânge și poate fi utilizat acolo ca precursor al testosteronului. Aproximativ 97% dintre androgeni se formează în testicule (aproximativ 7 mg / zi la bărbați), partea rămasă în glandele suprarenale . Androgenii acționează asupra epiteliului germinal și, legați de o proteină, sunt de asemenea transportați către celelalte organe țintă ale acestora prin sânge.
FSH se leagă de receptorii corespunzători de pe celulele Sertoli. Atât FSH cât și testosteronul controlează spermogeneza. FSH inițiază spermiogeneza, testosteronul promovează diviziunea celulelor mitotice și meiotice și astfel formarea spermatocitelor din spermatogonia, în timp ce FSH provoacă la rândul său maturarea finală a spermatidelor în spermă.
Suprimarea stimulării hormonale a funcției testiculare este în prezent intens cercetată în dezvoltarea contraceptivelor pentru bărbați . Testosteronul sau combinația acestuia cu antagoniști ai GnRH sau gestageni precum progestinul sunt în prezent considerați a fi cei mai promițători candidați. Acestea conduc la concentrații de testosteron mult reduse în testicul și astfel la o reducere puternică sau completă a formării spermei. În medicina veterinară, un preparat bazat pe analogul GnRH Deslorelin ( Suprelorin ® ) a fost aprobat din 2008 , care suprimă fertilitatea la bărbați timp de câteva luni. În plus, este aprobat un vaccin pentru porci ( Improvac ® ), care duce la formarea de anticorpi împotriva GnRH și astfel suprimă funcția testiculară.
Istoria dezvoltării

Condiția prealabilă pentru reproducerea sexuală este separarea celulelor specializate pentru reproducere (celule germinale) de celulele normale ale corpului (celule somatice). Această separare se realizează deja în ciliați sub forma unui micronucleu , mai clar decât în algele sferice , unde grupul celular principal al celulelor corpului este opus unui grup mic de celule germinale ( gonidia ), care, totuși, nu este încă dezvoltat sub forma unui organ delimitat . Prezența testiculelor (sau, în principiu, a gonadelor) nu este o caracteristică de bază a celulelor multicelulare.
În cazul bilateriei , apare pentru prima dată un al treilea cotiledon , mezodermul și, astfel , organele complexe. Cu toate acestea, la multe nevertebrate reproducerea sexuală este încă combinată cu posibilitatea reproducerii asexuale. Aici există adesea o schimbare de generație , adică cea sexuală urmează un ciclu de reproducere asexuat.
Diferențierea gonadelor în testicule și ovare este un semn distinctiv al speciilor segregate. Până în prezent nu s-a clarificat dacă hermafroditismul sau segregarea de gen sunt caracteristica plesiomorfă a bilateriei. Ambele gonade ies din același sistem în dezvoltarea embrionară. În multe grupuri de animale, în ciuda acestei separări de gen, este posibilă reproducerea fără fertilizare ( partenogeneză ), care poate fi privită ca o formă redusă de reproducere sexuală. Animalele masculine apar aici doar în cazuri excepționale. Partenogeneza se găsește la numeroși taxoni , de la rotifere la unele șopârle . Până la amfibieni, forme hibride sau o schimbare de gen ( dicogamie ) pot fi găsite și în timpul ontogenezei . Este posibilă transformarea ovarelor în testicule ( proteroginie ), precum și a testiculelor în ovare ( proterandrie ).
La majoritatea animalelor, se determină genetic dacă dispoziția inițială indiferentă de sex a gonadelor are ca rezultat un testicul sau un ovar . La viermi și muște, sexul este determinat de raportul dintre cromozomii X și autozomi . La insectele care formează starea, testiculele apar la descendenți din ouă nefertilizate și ovare la animale cu un set diploid de cromozomi, adică din ouă fertilizate. La mamifere, sexul este determinat de cromozomul Y . Pe acest cromozom sexual ( gonosom ) este localizată o genă ( regiunea determinantă a sexului Y , Sry ), care interacționează cu genele altor cromozomi și (la om începând cu săptămâna a 7-a după fertilizare) duce la formarea factorului determinant testicular . Aceasta inițiază dezvoltarea testiculelor și, astfel, a sexului masculin în general. Codifică o serie de factori de transcripție , așa-numitele proteine HMG ( proteine din grupul cu mobilitate ridicată ). Aceste proteine au numeroase alte funcții în alte țesuturi, iar mecanismele exacte implicate în formarea testiculelor sunt în prezent cercetate intens. Cu expresia Sry, celulele Sertoli se diferențiază, care, printre altele, produc hormonul anti-Müllerian și determină astfel regresia canalelor Müllerian . Dezvoltarea ulterioară a testiculului și a celorlalte caracteristici ale sexului masculin este controlată de androgeni. Cu toate acestea, în cazul unor grupuri de animale, sexul este determinat de factori de mediu. În cazul unor amfibieni și a multor reptile ( broaște țestoase , aligatoare ) , de exemplu, sexul depinde de temperatura de incubație. ( Vezi și. Determinarea sexului dependent de temperatură )
La multe animale, gonadele sunt dezvoltate în strânsă relație cu sistemul excretor ( rinichi primitiv , nefridie ), în special tractul urinar să fie împărțit ca sistem samenableitendes, motiv pentru care ambele sisteme de organe ale vertebratelor, precum și rezumatul aparatului urinar și sexual . Cu toate acestea, la nevertebrate, locul de origine, localizarea și canalele sunt foarte diferite, astfel încât se presupune că reproducerea sexuală a apărut de mai multe ori și independent una de cealaltă în evoluție . Complexitatea organelor genitale nu depinde de stadiul evoluției; este foarte mare la viermi plati, de exemplu.
Embriologia vertebratelor
Dezvoltarea testiculară la vertebrate
|

În embrion , testiculele și ovarele apar din același sistem , așa-numita creastă genitală . Se formează în zona rinichiului urnal și se extinde inițial de la torace până la coadă . La majoritatea vertebratelor, doar partea de mijloc a acestui sistem alungit devine gonada propriu-zisă, secțiunile rămase se dezvoltă în ligamentele goneale . Sub influența Sry, a celulelor germinale primordiale din sacul vitelin migrează în Anlage gonadal (la om , în a 6 săptămâni embrionare) și epiteliul cavitatea corpului primitiv ( coelom ) crește degetul asemănător în sistem , așa-numitul liniile germinale primare .
Cordele germinale sau testiculare pătrund în sistemul gonadal și cresc în jurul celulelor germinale primordiale. Gonadele sunt împărțite temporar în cortex și măduvă, deși la embrionii masculi doar măduva se dezvoltă în testicul, în timp ce cortexul se retrage. Procese similare au loc la indivizii genetic feminini, dar mai târziu și ovarul se formează din cortex în timp ce măduva degenerează. Conexiunea dintre cordoanele testiculare și suprafață se pierde în cele din urmă. Din corzile testiculare, prin intermediul celulelor pre-Sertoli care exprimă Sry , se dezvoltă celulele Sertoli , care sunt considerate organizatorii dezvoltării testiculare ulterioare și astfel interacționează cu celulele mioide . Spermatogonia provine din celulele sexuale primare .
În interior, corzile testiculare formează o rețea de corzi interconectate, rețeaua testiculară ulterioară ( rete testis ). Rețeaua testiculară se conectează la niște tubuli urnali care devin conductele eferente ale capului epididimului . Canalul urinar (canalul Wolff ) este, de asemenea, utilizat ca canal epididimal și canalul spermatic ca cale de drenaj a spermei. În lumenul tubulilor seminiferi nu se dezvolta pana la pubertate , la amfibieni după metamorfoză , până atunci cablurile testiculare sunt solide.
Tunica albuginea , cadrul țesutului conjunctiv al testiculului și celulele Leydig apar din partea mezodermică a anusului testicular . Celulele Leydig pot fi găsite și în dezvoltarea testiculară timpurie; ele exprimă factorul steroidogen 1 (Sf1), iar producția lor de testosteron determină în mod semnificativ dezvoltarea organelor sexuale masculine.
Îmbătrânire
La bărbați - spre deosebire de femei ( vezi menopauză ) - nu există o încetare bruscă a funcției gonadelor la o vârstă definită. Atât producția de hormoni, cât și maturarea celulelor germinale sunt potențial păstrate până la bătrânețe. De fapt, paternitatea a fost documentată până în deceniul zece al vieții . În realitate, însă, există diferențe individuale marcate și mulți bărbați devin mai devreme sau mai târziu infertili , ceea ce nu trebuie confundat cu impotența . Cauzele exacte ale acestor diferențe nu sunt cunoscute în detaliu; se suspectează factori vasculari , printre alții . Din punct de vedere statistic, din jurul celui de-al patrulea deceniu al vieții, involutia testiculelor , care progresează foarte lent, cu o scădere a greutății, dimensiunii și producției de spermă. Cu toate acestea, pentru individ, nu există nici o previziune de făcut.
Modificările structurale sunt , prin urmare , de asemenea , obiectul unei game largi, dar o imagine mixtă a valorilor normale și în mod clar atrofice tubii testiculare pot fi văzute ca tipic . O scădere semnificativă a producției de testosteron observată la unii bărbați poate duce la Climacterium virile cu bufeuri , cefalee și alte simptome .
Istoria explorării

În antichitate și Evul Mediu, testiculul era considerat doar o stație de tranzit pentru materialul seminal. Alkmaion von Kroton (începutul secolului al V-lea î.Hr.) a suspectat creierul ca fiind originea materialului seminal, care ajunge la testicule prin vasele de sânge . The atomists ( Anaxagora , Democrit ) și Aristotel au inclus măduva spinării în această cale, Galenus (125-199) , suspectat originea celulelor spermatice în vasele de sânge prin care acestea ajung la testicule. Aceste idei au persistat în Evul Mediu. Desenele anatomice ale lui Leonardo da Vinci arată conexiuni între testicul și plămâni și creier, deoarece da Vinci a suspectat originea „puterii spirituale” a materialului seminal din creier, în timp ce testiculele oferă doar baza materială pentru „impulsurile inferioare”.
Explorarea structurală
Abia odată cu începutul Iluminismului în secolul al XVII-lea, ideile materialului seminal masculin au fost demistificate și s-a recunoscut legătura directă dintre testicule și reproducere. Prima descriere modernă a structurii testiculului provine de la Reinier de Graaf (1641–1673). Nathaniel Highmore a descris în 1651 corpul țesutului conjunctiv al testiculului ( Corpus Highmori ), care a fost numit în 1830 de Astley Paston Cooper Mediastinum testis . În 1677, inventatorul microscopului , Antoni van Leeuwenhoek , a descoperit spermatozoizii, despre care credea că sunt organisme miniaturizate, pre-formate („animale de sămânță”).

Odată cu dezvoltarea tehnicilor histologice , structura fină a testiculului ar putea fi, de asemenea, elucidată. În 1841 anatomistul elvețian Albert von Koelliker a recunoscut pentru prima dată legătura directă dintre tubulii testiculari și spermatozoizi și a descoperit că celulele spermatozoizilor din acești tubuli apar ca produse de diferențiere celulară. În 1850, elevul lui Kölliker, Franz von Leydig, a descris pentru prima dată celulele intermediare ( celulele Leydig ).
În 1865 Enrico Sertoli a descoperit celulele de susținere ( celulele Sertoli ). În 1871 Victor Ebner a reușit să diferențieze celulele Sertoli de spermatogonia și cinci ani mai târziu La Valette St. George a inventat termenul „spermatogonia” și clasificarea etapelor individuale de dezvoltare a spermatozoizilor, care este și astăzi obișnuită. Celulele Sertoli au fost considerate mult timp ca sincitia , abia în 1956 Don W. Fawcett și Mario H. Burgos au reușit să demonstreze că fiecare celulă Sertoli are propriile limite ale celulelor.
Hugo Ribbert a recunoscut încă din 1904 că carminul administrat în sânge nu pătrunde în lumenul tubulilor seminiferi și în rețeaua testiculară. Multă vreme această descoperire a fost neglijată, deși a fost prima dovadă a barierei hemato-testiculare. Aceste cunoștințe au fost preluate abia la sfârșitul anilor 1950, iar în 1963 J. Brökelmann a reușit să demonstreze joncțiunile strânse ale celulelor Sertoli ca bază morfologică a barierei hemato-testiculare. Paul J. Gardner și Edward A. Holyoke au reușit să elucideze structura fină a barierei hemato-testiculare un an mai târziu.
Hormoni
Deși efectele castrării erau cunoscute de mii de ani, abia în 1849 Arnold Adolph Berthold a reușit să demonstreze experimental formarea hormonilor în testicule prin intermediul transplanturilor de testicule la cocoși. La o vârstă foarte înaintată, Charles-Édouard Brown-Séquard a întreprins autoexperimente la sfârșitul secolului al XIX-lea cu fluid din testiculele câinilor și cobai, pe care el i-a atribuit o putere de întinerire și întărire, deși era o cantitate mai mare de hormoni homeopati. că a obținut în acest fel. La începutul secolului al XX-lea, transplantul de testicule animale sub peretele abdominal a fost considerat un mijloc de întinerire, în special vienezii Robert Lichtenstern și Eugen Steinach au fost protagoniștii acestei metode. Steinach a dorit să realizeze acest proces de întinerire prin legarea canalului deferent (cel mai faimos pacient al său a fost Sigmund Freud ) și a descris transplantul testicular ca o „terapie” pentru homosexualitate . După 1945, aceste xenogrefe controversate s - au demodat.
În 1903, Pol Bouin și Paul Ancel au publicat pentru prima dată știința că celulele Leydig sunt locul în care sunt produși hormoni sexuali masculini. În 1931, Adolf Butenandt și Kurt Tscherning au izolat androsteronul (un metabolit al testosteronului) din urina bărbaților, în 1935 Ernst Laqueur a reușit să izoleze el însuși testosteronul din testiculele de taur și a inventat și numele acestui hormon (din testicul „testicul” și „ steroid ").
Existența hormonilor nesteroidieni în testicule a fost postulată încă din anii 1920, dar abia în 1932 D. Roy McCullagh și-a demonstrat funcționalitatea la șobolani castrați și l-a numit inhibină . Deși bioanalizele acestui hormon au fost dezvoltate în anii 1960 , existența sa a fost controversată timp de câteva decenii și nu a fost acceptată pe scară largă până în 1979. În 1984/85 au fost relevate structura și subtipurile hormonului. Odată cu clarificarea secvenței ADN care codifică hormonul , apartenența inhibinei la grupul factorilor de creștere care transformă β a fost recunoscută în 1985 .
În timp ce relațiile dintre LH și testosteron erau deja cunoscute în anii 1960, dependența de FSH a celulelor Sertoli a fost dovedită abia în 1984 de Joanne M. Orth .
Diferențierea de gen
Baza cromozomială a diferențierii de sex a fost deja clarificată între 1910 și 1916, în principal prin lucrarea lui Thomas Hunt Morgan privind muștele fructelor , pentru care a primit Premiul Nobel pentru fiziologie sau medicină din 1933 . Alfred Jost a recunoscut în 1947 că gonadele sunt determinate în primul rând de sexul feminin și că sexul masculin este dependent de testosteron. Cu toate acestea, abia la începutul anilor 1960, cromozomul Y a fost identificat ca un factor major la mamifere. Localizarea exactă a genei pentru factorul determinant testicular nu a fost determinată până în 1990; diversele sale funcții nu sunt încă cunoscute în detaliu și fac obiectul cercetărilor actuale.
Tulburări de dezvoltare și boli
Leziunile testiculului apar la oameni ca traume contondente , în special în artele marțiale și luptele. Există riscul de sângerare sub capsula testiculară ( hematocel ), care de obicei necesită tratament chirurgical. Leziunile cu deschiderea scrotului (puncția și rănile prin împingere, la animale mușcături, sârmă ghimpată etc.) pot provoca inflamații testiculare (vezi mai jos) sau chiar abcese , precum și peritonită datorită conexiunii deschise a procesului vaginal cu cavitatea abdominală .
Malformații
O anorhie este absența ambelor testicule; dacă se formează un singur testicul, se vorbește despre monarhie . Aproximativ 5% dintre copiii bărbați operați pentru că nu au coborât testiculele au doar unul sau niciun testicul. Au adesea mici noduri de țesut conjunctiv cu celule Leydig împrăștiate. Deoarece un testicul funcțional intact este absolut necesar pentru exprimarea genului masculin, cel puțin un testicul intact trebuie să fi fost prezent în faza embrionară, care poate să fi regresat mai târziu.
În cazuri rare, ca urmare a tulburărilor de dezvoltare, ovarele pot apărea și pe lângă testicule la om și la alte mamifere ( hermafroditism verus - hermafrodite „reale”; vezi și intersexualitatea ). Cu anumite malformații ale gonadelor ( disgeneză gonadică ), testiculele nu sunt create, rămân subdezvoltate sau conțin țesut ovarian ( ovotestis ).
O dezvoltare nedorită foarte rară este fuziunea splenogonadală cu o legătură între țesutul testicular și splină .
Cu tulburări ale migrației testiculului ( Maldescensus testis ) pot apărea diferite anomalii de poziție . Testiculul poate rămâne în cavitatea abdominală ( criptorhidie , „ testicul abdominal”), poate rămâne blocat în canalul inghinal („testicul inghinal”, „osul alunecător”) sau poate lua o cale greșită și, de exemplu, ajunge să se întindă sub piele a inghinei sau a coapsei interioare ( ectopia testiculară ). Testicul Maldescensus este una dintre cele mai frecvente malformații la om și apare la 3 până la 5% dintre nou-născuți și 33% dintre bebelușii prematuri; criptorhidele apar, de asemenea , cu o frecvență similară la animalele domestice și duc la excluderea din reproducere. Dacă testiculul este poziționat incorect, spermatozoizii fertili nu se pot forma din cauza sensibilității la temperatură a epiteliului germinal, dar producția de androgeni este menținută. Nerespectarea coborârii mai mult de doi ani poate duce la pierderea spermatogoniei și, prin urmare, la modificări ireversibile ale testiculului.
Statura scurtă ereditară a testiculelor ( hipoplazie testiculară ) este relativ frecventă la animalele de companie. Hipoplazia testiculară poate fi cauzată și de tulburări cromozomiale ( sindromul Klinefelter ), infecții sau tulburări hormonale .
O mărire anormală a testiculelor se numește macroorhidie .
Inflamația testiculelor
Inflamația testiculelor ( orhita ) poate apărea cu leziuni ale scrotului cu invazia bacteriilor sau cu unele boli infecțioase. Orhita este o posibilă complicație a oreionului , a infecțiilor cu virusul Coxsackie și a varicelei la om . De asemenea, bruceloză și tuberculoză se pot manifesta pe testicule. La animale, de asemenea, tuberculoza și poate bruceloză și pseudotuberculoză ( oaie ), anemia infecțioasă ecvină și anemia infecțioasă felină ( pisici ) pot fi însoțite de o orhidită. Inflamația testiculelor poate duce la micșorarea testiculelor ( atrofie testiculară ) și infertilitate , deoarece nu se mai pot forma celule de spermă ( aspermie ) sau spermatozoizi care funcționează.
Tulburări circulatorii
Ca varicocel se numește varice, extensii similare, care se referă în special la venele din stânga ale plexului pampiniform în cordonul spermatic. Un varicocel poate duce la producția limitată de spermă în testicul pe aceeași parte.
Acumularea de lichid seros în tecile testiculare se numește o hidrocel sau fractură de apă. În plus, fluidul se poate colecta și în cordonul spermatic, care este denumit apoi hidrocel funiculi spermatici .
O torsiune este o rotație anormală a testiculului, în care prinderea elicoidală a cordonului spermatic și a venelor de drenare poate duce la moartea testiculului. Torsiunea severă a testiculelor este o urgență foarte dureroasă și se poate aștepta la deteriorarea permanentă a testiculului după doar două ore. Anexa testiculului se poate dezvolta , de asemenea , un așa-numitul hidrat de torsiune.
Tulburări circulatorii cu risc de dezvoltare a necrozei sunt observate și în afecțiunile vaselor de sânge, cum ar fi purpura Henoch-Schönlein , endangiita obliterantă și panarterita nodoză la om, arterita la cai și, în general, în tromboză .
Tumori
O tumoare testiculară este o mărire patologică a testiculului. Tumorile testiculare pot fi benigne sau maligne .
Majoritatea inofensivelor măriri testiculare sunt chisturi . Două tipuri diferite de chisturi se pot dezvolta pe testicul. Hidrocelele sunt proeminențe din tunica vaginală a testiculului care conțin un lichid limpede, de culoare chihlimbar. Acestea sunt cauzate de leziuni sau inflamații. Spermatocelii provin din rete testicul sau epididim și conțin spermă. Teratoamele sunt în mare parte tumori benigne ale celulelor germinale.
O cauză foarte rară a unei mase pe testicul poate fi fuziunea splenogonadică .
Tumorile testiculare maligne ( cancerul testicular ) sunt subdivizate în degenerări ale celulelor germinale ( tumori testiculare germinale : seminome ) și non- seminome . Degenerarea celulelor germinale este cel mai frecvent cancer la bărbații cu vârste cuprinse între 20 și 40 de ani și reprezintă aproximativ 90% din toate tumorile testiculare. Principalul factor de risc sunt testicule care nu au migrat in scrot. Restul de 10% se datorează degenerare tumorous a altor părți de țesut ( tumoare Sertoli de celule , celule tumorale Leydig , limfom non-Hodgkin, etc.).
Defecțiuni
În plus față de bolile menționate mai sus , pot apărea și substanțe chimice care traversează bariera sângelui-testiculului, cum ar fi toxinele din mediu (de exemplu, cadmiu ), aditivi la materialele de ambalare (de exemplu , ftalați , dietilhexil adipat ), unele medicamente (de exemplu furazolidonă ) și hormoni ( vezi și perturbatoare endocrine ) sau radiațiile ionizante pot duce la afectarea severă a epiteliului tubilor seminiferi. Deoarece formarea spermei este asociată cu rate foarte mari de diviziune celulară ( mitoză , meioză ), epiteliul germinal este deosebit de sensibil la toxinele celulare. Astfel de daune pot duce la modificări multiple până la absența completă a spermei ( vezi și analiza spermei ).
Producția inadecvată de androgeni este cunoscută sub numele de hipogonadism . Aceasta poate fi congenitală, secundară bolilor testiculului sau datorită deficitului de gonadotropină (de exemplu, hipofuncția glandei pituitare , sindromul olfactogenital ).
Criptorhidia abdominală la o oaie domestică („rinichi”)

Hipoplazie testiculară la pisică, în dreapta testicul dezvoltat în mod normal.

Varicocel distinct pe stânga.
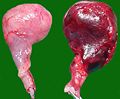
Testiculele câinilor eliminate după rotație (stânga: organ sănătos)
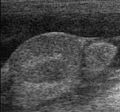
Sonografie : câine cu criptorhidie abdominală și ascită (stânga: testicule, dreapta: epididim)





examinare
Examinarea testiculara este un bază importantă de examinare pentru oameni și animale , cu un scrot. Aici se verifică prezența, dimensiunea, localizarea și consistența testiculului. Examenul cu ultrasunete este utilizat în principal ca metodă de imagistică . Volumul testicular este determinat fie prin comparație cu așa-numitul lanț Prader, fie prin măsurarea cu ultrasunete. Diafanoscopia are încă o anumită valoare în diagnosticul hidrocelului . O biopsie testiculară poate fi efectuată pentru prelevarea de probe de țesut . La animalele cu testicule localizate în cavitatea abdominală, endoscopia este utilizată în primul rând pe lângă examinarea cu ultrasunete .
O examinare funcțională este crearea unei analize a materialului seminal . Aici se evaluează numărul, forma și mobilitatea spermatozoizilor. Determinarea nivelului de inhibină B în sânge este utilizată ca marker pentru funcția și fertilitatea celulei Sertoli, dar valoarea sa informativă este controversată.
Testul de stimulare a celulelor Leydig poate fi utilizat pentru a detecta țesutul testicular care nu poate fi detectat folosind tehnici de imagistică .
castrare

Restricția funcției testiculare se numește castrare. Se poate face prin îndepărtarea chirurgicală a testiculului ( orhidectomie ), ligarea vaselor testiculare („castrare fără sânge”), radiații sau substanțe chimice. Castrările se efectuează în principal la oameni pentru cancerul testicular. Testiculele îndepărtate chirurgical sunt de obicei înlocuite de o proteză testiculară din motive cosmetice .
Ca simbol al neputernicirii, castrarea joacă, de asemenea, un rol în mitologia multor culturi (vezi și teama de castrare ). În mitologia egipteană, Horus smulge testiculele de la adversarul său Seth . În mitologia greacă , Kronos îndepărtează mai întâi testiculele tatălui său Uranus și este ulterior emasculat chiar de fiul său Zeus . Pentru a renunța lumești dorește, auto- masculinizare a Galloi (preoți) în Cybele cultul frigieni , care , de asemenea , răspândit în Grecia antică și Roma, a fost comună, așa cum a fost cazul cu hijras din India . Cu toate acestea, în iudaism, castrarea, atât a oamenilor, cât și a animalelor, este strict interzisă. În creștinism , castrarea a fost, de asemenea, mal văzută. Eunucilor nu li s-a permis să fie hirotoniți preoți, dar existau curenți în care autocastrarea era efectuată ca ritual (vezi Skopzen ).
Din punct de vedere istoric, sclavii , prizonierii de război, cântăreții sau gărzile haremurilor (vezi Palace Eunuch ) au fost de asemenea castrate. Castrarea non-justificată din punct de vedere medical a vizat în special prevenirea caracteristicilor sexuale secundare (tonul vocii, comportamentul sexual) cauzate de testosteron. Castrati au fost populare în viața muzicală europeană în 17 și 18 secole și au fost de multe ori a avut loc în ceea ce privește ridicat. Senesino , Farinelli , Caffarelli și Antonio Bernacchi sunt printre cei mai renumiți castrati din secolul al XVIII-lea .
Bărbații castrați nu se pot reproduce singuri. La fel ca clericii care s-au abținut voluntar , aceștia au fost considerați mai fiabili și folosiți ca funcționari și servitori în diferite societăți. Castrarea voluntară a infractorilor sexuali este încă o metodă de terapie, deși una controversată, în Germania și în unele state ale SUA.
În medicina veterinară, castrarea este utilizată, pe lângă indicațiile medicale (cancer testicular, boli ale prostatei și ale glandelor anale ), mai ales pentru a evita descendenții, pentru a îmbunătăți manevrabilitatea animalelor de companie ( castrat , bou ), pentru a crește performanța de îngrășare și calitatea cărnii, la porcii domestici, de asemenea, pentru a evita „mirosul de mistreț” al cărnii efectuate. Castrările pe animale s-au efectuat probabil încă de la începutul neoliticului . Castrarea este una dintre puținele îndepărtări de organe care nu sunt indicate din punct de vedere medical și care sunt permise și astăzi în Germania în temeiul Legii privind bunăstarea animalelor (§ 6), la animale foarte tinere chiar și fără eliminarea durerii, care, totuși, nu este de necontestat.
Semnificație culturală și istorică

În mitologia japoneză , tanuki , demonii ( yōkai ) asemănători câinelui raton , sunt adesea reprezentați ca un simbol al norocului cu testiculele supradimensionate. În Grecia antică , organele genitale ale animalelor, în special taurii ( taurobolium ), erau oferite ca sacrificii. Testiculele au fost considerate un simbol al virilității și al potenței creative. Potrivit lui Taylor, ei aveau o putere simbolică mai puternică decât penisul până la sfârșitul secolului al XVI-lea .
botanică

Se consideră că plantele, care seamănă cu organele genitale masculine, au un efect afrodisiac și de creștere a fertilității. Filosoful grec Theophrastus von Eresos le-a numit Orchis , denumirea greacă pentru testicule, datorită asemănării celor două bulbi ai orhideei cu testiculele. Se spune că consumul lor este favorabil nașterii unui băiat („orhidee”). Ulterior Orchis a dat numele întregii familii de orhidee .
Numele de avocado este derivat din cuvântul indian ahuacatl (testicul), care se referă la forma de testicul a fructului acestui copac.
În art
În artă, spre deosebire de falus , testiculele nu joacă un rol central în afara eroticii și a pornografiei . „Sângele și testiculele”, un joc fonologic de cuvinte despre ideologia sângelui și a solului , este adesea folosit în mod disprețuitor în critica de artă .
Unul dintre personajele din Tristan al lui Thomas Mann este domnul Klöterjahn („Klöten” este expresia low-germană pentru testicule, un simbol al aptitudinii pentru viață și vitalitate). În romanul din 2002 Sanningen om Sascha Knisch ( eng . Titlul adevărul despre Sascha Knisch. 2003) al autorului suedez Aris Fioretos , testiculele sunt laitmotivul . Filmul german Eierdiebe tratează subiectul cancerului testicular și al pierderii unui testicul.

Stema casei nobile italiene Colleoni din Bergamo prezintă mai multe perechi de testicule și este probabil o aluzie la coglione , un nume italian pentru testicule. O statuie a lui Bartolomeo Colleoni cu această stemă pe bază se roagă personajul principal din primul volum al romanului în trei părți al lui Heinrich Mann The Goddesses or The Duchess of Assy Three Novels .
Ca mâncare

Testiculele sunt procesate ca alimente în multe regiuni. Germania a fost singura țară din UE în care testiculele au fost interzise ca alimente. Conform Regulamentului UE nr.853 / 2004 din 29 aprilie 2004, testiculele sunt singurele organe genitale ca hrană, toate celelalte sunt clasificate ca nepotrivite pentru consum ( confiscă ).
diverse
- Americanul Gregg Miller a primit premiul Ig Nobel pentru medicină în 2005 pentru dezvoltarea protezelor testiculare pentru câinii castrați .
- În martie 2006, Volkswagen promovase Golf GTI pe afișe cu „Turbo Cojones” în SUA . În engleză, termenul cojones înseamnă curaj și îndrăzneală, în spaniolă, unde cojones înseamnă „testicule” și colocvial „tăiere” sau „curaj”, combinația de cuvânt înseamnă literalmente „turbo testicule”. Promovarea a fost retrasă după proteste.
- Glandele în formă de pungă, de dimensiuni de ouă de găină (saci de ricin) sub osul pubian al castorului erau cunoscute și sub denumirea de „testicule” în trecut.
literatură
- AJP van den Brock: Gonade și modalități de execuție. În: Bolk și colab. (Ed.): Manual de anatomie comparativă a vertebratelor . Volumul 6, Urban & Schwarzenberg, Berlin 1933, pp. 1–154.
- W. Busch, A. Holzmann (Ed.): Andrologie veterinară. Schattauer, Stuttgart 2001, ISBN 3-7945-1955-8 .
- U. Gille: organe de reproducere masculine. În: F.-V. Salomon și colab. (Ed.): Anatomie pentru medicina veterinară . Enke, Stuttgart 2004, ISBN 3-8304-1007-7 , pp. 389-403.
- R. Hautmann, H. Huland: Urologie . Springer, Berlin 2006, ISBN 3-540-29923-8 .
- H.-G. Liebich: histologie funcțională a mamiferelor domestice. Ediția a IV-a. Schattauer, Stuttgart 2003, ISBN 3-7945-2311-3 .
- JD Neill (Ed.): Fiziologia reproducerii Knobil și Neill. 3. Ediție. Academic Press, Amsterdam 2005, ISBN 0-12-515400-3 .
- PE Petride: Funcții endocrine IV. Sistem hipotalamo-hipofizar și țesuturi țintă. În: G. Löffler, PE Petrides (Ed.): Biochimie și patobiochimie. Ediția a VII-a, Springer, Berlin 2003, ISBN 3-540-42295-1 , pp. 865-908.
- U.N. Riede și colab.: Sistemul genital masculin. În: U.-N. Riede și colab. (Ed.): Patologie generală și specială . Thieme, Stuttgart 1989, ISBN 3-13-683302-3 , pp. 768-779.
- B. Vié: Testicule. Fête de paires, mythology, les lingerie, d'une curiosité culinaire, les attributs du sujet, lexique. Edition de l'Epure, Paris 2005, ISBN 2-914480-58-X (numeroase rețete, îmbogățite cu informații culturale și istorice).
- R. Wehner, W. Gehring: Zoologie. Ediția a 23-a, Thieme, Stuttgart 1995, ISBN 3-13-367423-4 .
- U. Welsch: histologie manuală Sobotta. Urban & Fischer, München 2002, ISBN 3-437-42420-3 .
Link-uri web
Dovezi individuale
- ↑ Walther Graumann: Anatomie compactă a manualului. Volumul 3, Schattauer, Stuttgart 2004, ISBN 3-7945-2063-7 , p. 265.
- ↑ H. Sosnik: Studii privind dimensiunea gonadei masculine umane în biomorfoză, intoxicație cu alcool și ciroză - o analiză și descoperiri proprii. În: Anuarul morfologic al lui Gegenbaur. Vol. 134, nr. 5, 1988, pp. 733-761, PMID 3224804 .
- ^ David AE Spalding: Balenele de pe coasta de vest . Editura Harbour, 1999, ISBN 1-55017-199-2 .
- ↑ I. Gerendai u a.: Inervația și receptorii serotoninergici ai testiculului interacționează cu acțiunea locală a interleukinei-1beta asupra steroidogenezei. În: Neuroștiințe autonome. 7 iulie 2006, PMID 16829209 .
- ↑ I. Ducic, AL Dellon: Durerea testiculară după repararea herniei inghinale: o abordare a rezecției ramurii genitale a nervului genitofemoral. În: Jurnalul Colegiului American al Chirurgilor. Volumul 198, nr. 2, februarie 2004, pp. 181-184, PMID 14759772 .
- ↑ FD Brown și colab.: Organul ofertantului în broasca Bufo marinus: Efectele orhidectomiei asupra morfologiei și expresiei polipeptidei asociate lamelelor 2. În: Dezvoltarea și diferențierea dezvoltării. Volumul 44, nr. 6, decembrie 2002, pp. 527-535, PMID 12492511 .
- ↑ CF Farias: Organul ofertantului Bufo ictericus: o analiză microscopică cu lumină și electronică. În: Micron. Volumul 33, nr. 7-8, 2002, pp. 673-679, PMID 12475564 .
- ↑ G. Michel: Sistemul sexual. În: F.-V. Salomon (ed.): Manual de anatomie a păsărilor . Fischer-Verlag, Stuttgart 1993, ISBN 3-334-60403-9 , pp. 197-226.
- ^ RP Amann, SS Howards: Producția zilnică de spermatozoizi și rezervele de spermatozoizi epididimali la masculul uman. În: Journal of Urology . Vol. 124, nr. 2, august 1980, pp. 211-215, PMID 6772801 .
- ↑ C. Petersen, O. Soder: Celula sertoli - o țintă hormonală și „super” asistentă pentru celulele germinale care determină mărimea testiculului. În: Hormone Research Volumul 66, nr. 4, 2006, pp. 153-161, PMID 16804315 (text integral) .
- ↑ M. Fijak, A. Meinhardt: Testicul în privilegiul imunitar. În: Imunologic Review-uri . Volumul 213, octombrie 2006, pp. 66-81, PMID 16972897 .
- ↑ RS Swerdloff și colab.: Suprimarea spermatogenezei la om indusă de antagonistul hormonal care eliberează gonadotropina Nal-Glu și enantatul de testosteron (TE) este menținută doar de TE. În: Journal of Clinical Endocrinology and Metabolism . Volumul 83, nr. 10, octombrie 1998, pp. 3527-3533, PMID 9768659 .
- ↑ KL Matthiesson: Efectele testosteronului și levonorgestrelului combinate cu un inhibitor de 5alfa-reductază sau antagonist al hormonului care eliberează gonadotropina asupra spermatogenezei și a nivelurilor de steroizi intratesticulari la bărbații normali. În: Journal of Clinical Endocrinology and Metabolism . Vol. 90, nr. 10, octombrie 2005, pp. 5647-5655, PMID 16030154 .
- ↑ Peter Y. Liu și colab.: Determinanți ai ratei și extinderii suprimării spermatogene în timpul contracepției hormonale masculine: o analiză integrată. În: Journal of Clinical Endocrinology and Metabolism. Volumul 93, nr. 5, pp. 1774-1783, doi: 10.1210 / jc.2007-2768 .
- ↑ Improvac la vetpharm.uzh.ch
- ↑ a b c d C. Tilmann, B. Capel: Căi celulare și moleculare care reglementează determinarea sexului mamiferelor. În: Progrese recente în cercetarea hormonală. Volumul 57, 2002, pp. 1-18.
- ↑ C. Dournon și colab.: Inversarea temperaturii la amfibieni și reptile. În: International Journal of Dev Biology. Volumul 34, numărul 1 martie 1990, pp. 81-92. PMID 2393628
- ^ E. Nieschlag și colab.: Funcțiile de reproducere la tinerii și bunicii tineri. În: Journal of Clinical Endocrinology and Metabolism . Vol. 55, 1982, pp. 676-681.
- ↑ H. Bürgi, C. Hedinger: Modificări testiculare histologice la bătrânețe. În: săptămânal medical elvețian. Volumul 47, 1959, pp. 1236-1239.
- ↑ A. von Kolliker: Contribuții la cunoașterea relațiilor sexuale și a fluidului seminal al nevertebratelor, împreună cu un experiment despre natura și semnificația așa-numitelor animale de sămânță . Berlin 1841.
- ↑ AHJ La Valette St. George: Despre geneza corpurilor semințelor. În: Arhive pentru anatomie microscopică. Volumul 12, 1876, pp. 797-825.
- ^ J. Brökelmann: Structura fină a celulelor germinale și a celulelor Sertoli în timpul ciclului epiteliului seminifer la șobolan. În: Cercetare celulară și anatomie microscopică. Vol. 59, 1963, pp. 820-850, PMID 14015736 .
- ^ PJ Gardner, EA Holyoke: Structură fină a tubului seminifer al șoarecelui elvețian. I. Membrana limitativă, celula Sertoli, spermatogonia și spermatocitele. În: Înregistrare anatomică . Volumul 150, decembrie 1964, pp. 391-404, PMID 14248309 .
- ↑ AA Berthold: Transplantul testiculelor. În: Arhive pentru anatomie și fiziologie. Departamentul fiziologic . Volumul 16, 1849, pp. 42-46.
- ↑ F. Mildenberger: întinerire și „vindecare” a homosexualității. Eugen Steinach la vremea lui. În: Cercetare sexuală. Volumul 15, 2002, pp. 302-322.
- ^ P. Bouin, P. Ancel: Recherches sur les Cellules interstitielles du testicule chez les mammifères. În: Archives de zoologie expérimentale et générale. Seria 4, Volumul 1, 1903, pp. 437-523.
- ↑ K. David și colab.: Despre hormonul cristalin din testicule (testosteron). În: Jurnalul de chimie fiziologică . Volumul 233, 1935, pp. 281-282.
- ↑ A. Lampel: Comentariu. În: Urologie curentă. Volumul 35, 2004, pp. 6-8. (Text integral) ( Memento din 14 noiembrie 2007 în Arhiva Internet )
- ↑ Axel Wehrend: simptome cheie de ginecologie și obstetrică la câini. Enke, Stuttgart 2010, ISBN 978-3-8304-1076-8 , p. 57.
- ^ A b c G. Taylor: Castration: An Abbreviated History of Western Manhood. Routledge, 2002, ISBN 0-415-93881-3 .
- ↑ W. Bittorf: sânge și testicule . În: Der Spiegel . Nu. 44 , 1976, pp. 228-232 ( Online - 25 octombrie 1976 ).
- ^ Peter Philipp Riedl: Imagini de epocă - tipologii de artiști. Contribuții la versiunile tradiționale în literatură și știință din 1860 până în 1930. Vittorio Klostermann, 2005, ISBN 3-465-03410-4 , ( p. 570 ).
- ↑ Regulamentul (CE) nr. 853/2004 al Parlamentului European și al Consiliului. ( europa.eu )
- ↑ Miriam Jordan: Bun gust pierdut în traducerea anunțului . Pe: articles.sun-sentinel.com din 19 martie 2006 (engleză).
- ↑ „Turbo-Cojones”, esloganul Volkswagen care moleste în EEUU . Pe: elmundo.es din 21 martie 2006 (spaniolă).
- ↑ Fotografia afișului. Pe: secure.flickr.com din 20 martie 2006.
- ↑ Dieter Lehmann: Două cărți de prescripție medicală din secolul al XV-lea din Rinul superior . Partea I: Text și glosar . Horst Wellm, Pattensen / Han. 1985; acum (= cercetare istorică medicală Würzburg. Volumul 34). Königshausen & Neumann, Würzburg, ISBN 3-921456-63-0 , p. 194.